
Open Journal of
Clinical and Medical Images
Research Article - Open Access, Volume 4
Molecular characterization of breast cancer cell lines in correlation with clinical markers
Dharambir Kashyap; Huda Salman*
Division of Hematology and Oncology, School of Medicine, Indiana University, Brown Center for Immunotherapy, Melvin and Bren Simon Comprehensive Cancer Center, Indianapolis, IN 46202, USA.
*Corresponding Author: Huda Salman
Department of Medicine, Professor of Immunotherapy
Professor of Medicine, Brown Center for Immunotherapy
Don Brown 975 W. Walnut St., IB554A, Indianapolis, IN
46202, USA.
Email: hsalman@iu.edu
Received : Apr 30, 2024
Accepted : May 24, 2024
Published : May 31, 2024
Archived : www.jclinmedimages.org
Copyright : © Salman H (2024).
Abstract
Cell lines are indispensable for biomedical research, offering a consistent platform for investigating cellular processes and disease mechanisms. Despite their value, cell lines possess limitations such as genetic drift and phenotypic alterations over successive passages, potentially compromising their relevance. Monoclonal derivation may not represent in vivo heterogeneity, particularly in cancer. Therefore, characterizing them becomes imperative. We aimed to identify breast cancer subtype-specific cell lines by assessing ER, PR, and HER-2 expression via immunohistochemistry and their concordance with molecular expression. We selected cell lines with consistent phenotypes that mimic primary cells and confirmed concordance in ER, PR, and HER-2 expression at protein and mRNA levels. Evaluation of ER, PR, and HER-2 was conducted using Immunocytochemistry (ICC), Immunofluorescence (IF), and real-time PCR across commonly used breast cancer cell lines. Results revealed differential expression patterns, with MCF-7 and T-47D cells showing positive ER and PR staining but negative HER-2 staining, while SK-BR-3 and HCC1954 cells exhibited positive HER-2 staining and negative ER and PR staining. Conversely, MDA-MB-231 and MDA-MB-468 cells displayed negative staining for all three markers. Real-time PCR analysis highlighted variations in HER-2 expression among the cell lines. Our investigation underscores MCF-7, SK-BR-3, MDA-MB-231, and MDA-MB-468 cell lines as pertinent models for studying major molecular subtypes of breast cancer.
Citation: Kashyap D, Salman H. Molecular characterization of breast cancer cell lines in correlation with clinical markers. Open J Clin Med Images. 2024; 4(1): 1186.
Introduction
Breast cancer stands as the foremost contributor to female cancer-related fatalities on a global scale [1]. Through gene expression profiling, breast cancer has been stratified into four distinct molecular subtypes. These include Luminal A and Luminal B, characterized by positivity for Estrogen Receptor (ER) and/or Progesterone Receptor (PR); Human Epidermal Growth Factor 2 (HER-2) enriched subtype, distinguished by the overexpression or gene amplification of the HER-2/neu gene; and triple-negative breast cancer, which lacks expression of ER, PR, and HER-2 [2]. Immunohistochemistry (IHC) is an essential technique utilized for the stratification of breast cancer into distinct molecular subtypes based on the staining patterns of Estrogen Receptor (ER), Progesterone Receptor (PR), and Human Epidermal growth factor 2 (HER-2) Receptors [3]. The existence of these subtypes underscores the heterogeneity of breast cancer, with patients harboring different subtypes exhibiting diverse genetic profiles and clinical outcomes [3,4]. In vitro studies have contributed substantially to the understanding of breast cancer heterogeneity. Therefore, it is imperative to ascertain whether cell lines retain the molecular characteristics of subtypes observed clinically [5]. While cell lines serve as representative models of breast cancer features to a considerable extent, reports have identified genetic and epigenetic alterations during initial maintenance and subsequent passaging [6]. Several studies have classified breast cancer cell lines into distinct molecular groups, revealing their comprehensive nature [7-9]. For instance, one study categorized cell lines into luminal, basallike, and mesenchymal subtypes [10]. Another study, utilizing gene expression profiling, delineated five subtypes among 51 cell lines: luminal, luminal/HER-2 positive, ER-negative/HER-2 positive, basal-like, and normal-like [5]. Triple-negative breast cancer has been further subclassified into basal-like 1, basal-like 2, immunomodulatory, mesenchymal, mesenchymal stem-like, and luminal androgen receptor subtypes [11]. Notably, MCF-7, T-47D, SK-BR-3, and MDA-MB-231 are preferred cell lines for breast cancer studies, collectively representing over two-thirds of cell lines utilized. Hence, it is pertinent to investigate the extent to which these cell lines mirror or diverge from clinical breast tumor subtypes. We aimed to characterize a panel of breast cancer cell lines of different molecular subtypes using markers commonly employed in research. This characterization may empower researchers to make more informed decisions when selecting cell lines representative of specific breast cancer molecular subtypes.
Methods
Cell line: Breast cancer cell lines MCF-7, T-47D, SK-BR-3, HCC1954, MDA-MB-231, MDA-MB-468, and MCF-10A, were purchased from ATCC. All cell lines were maintained in DMEM, high glucose media (Thermofisher Waltham, MA USA, #11965092) supplemented with 10% Fetal Bovine Serum (Thermofisher Waltham, MA USA, #26140079) and 1% antibiotic-antimycotic solution (Thermofisher Waltham, MA USA, #15240062), incubated at 37oC under constant supply of 5% CO2
Immunocytochemistry (ICC): For all used cell lines, 5x103 cells were grown on a 20 mm glass coverslip placed inside a 24-well plate. After 24 hours cells were washed gently with 1X PBS twice and fixed with 4% formaldehyde by incubating at room temperature for 15 minutes. Cells were permeabilized with 0.1% X-100 Triton at room temperature for 15 minutes and endogenous blocking was done with 1% BSA at room temperature for 45 minutes. Then each cell line was incubated with anti-estrogen receptor (abcam, Waltham, MA, USA, #ab16660), anti-progesterone receptor (abcam, Waltham, MA, USA, #ab101688), and anti-Her-2 positive (abcam, Waltham, MA, USA, #ab214275); primary antibodies in separate wells for overnight at 4OC (see table 1). The next day, the cells were incubated with Goat anti-Rabbit IgG (H+L) Secondary Antibody, HRP (Thermofisher Waltham, MA USA, #31460) for 60 minutes at room temperature. A secondary antibody was detected with DAB Quanto (Epredia Netherlands, B.V., #TA-060-QHDX) chromogen treatment for 5 minutes. Counterstaining was done with vector hematoxylin (#H-3401). Then the glass cover slip was taken out from the well and mounted on a glass slide with ProLong™ Glass Antifade Mountant (Thermofisher Waltham, MA USA, #P36984).
Immunofluorescence (IF): For all cell lines used, 5x103 cells were grown on a 20 mm glass coverslip placed inside a 24-well plate. After 24 hours cells were washed gently with 1X PBS twice and fixed with 4% formaldehyde by incubating at room temperature for 15 minutes. Permeabilized with 0.1% X-100 Triton at room temperature for 15 minutes. Endogenous blocking was done with 1% BSA at room temperature for 45 minutes. Then each cell line was incubated with anti-estrogen receptor (abcam, Waltham, MA, USA, #ab16660), anti-progesterone receptor (abcam, Waltham, MA, USA, #ab101688), and anti-Her-2 positive (abcam, Waltham, MA, USA, #ab214275); primary antibodies in separate wells for overnight at 4OC (Table 1). Next day, incubated with Alexa Flour 555 goat anti-rabbit IgG (Thermofisher Waltham, MA USA, #A-21428) secondary antibody for 60 minutes at room temperature. Fluorescent phalloidin iFluor 488 (abcam, Waltham, MA, USA, #ab176753) reagent was used for cytoskeleton counterstaining. Counterstaining was done with DAPI (Invitrogen, USA, #D1306). Then the glass cover slip was taken out from the well and mounted on a glass slide with ProLong™ Glass Antifade Mountant (Thermofisher Waltham, MA USA, #P36984). The cells were washed with 1X PBS in between after each step.
Quantification ICC & IF: ER and PR staining were scored according to nuclear staining intensity, weak, moderate, and high, and extension through the following criteria: 0%, negative; 1 to 25%, low expression; 26 to 50%, moderate expression; 51 to 75%, high expression; above >75%, very high expression. HER2 expression was analyzed according to the American Society of Clinical Oncology (ASCO)/College of American Pathologists (CAP) HER-2 test guidelines.
Nucleic acid isolation: Total RNA was isolated from each cell line (2x106 cells) using the ReliaPrep RNA cell miniprep system (Promega, #Z6011) following manufacturer instructions. Genomic DNA was also isolated from each cell line (2x106 cells) using QIAamp mini-Kit (Qiagen, #51104) following manufacturer instructions with slight modifications.
Real-time PCR: Real-Time PCR was performed for relative quantification of ESR1, ESR2, ESRα36, ESRα66, PGR, ERBB2, and AR genes with cDNA synthesized with 1 µg of total RNA using iScript cDNA synthesis kit (Bio-rad, Hercules, CA, USA, #1708890) and for ERBB2 gene with gDNA (50 ng). In both cases, the realtime reaction was performed with iTaq Universal SYBR green supermix (Bio-rad, Hercules, CA, USA, #172-5120) on QuantStudio Real-Time PCR systems (Thermofisher Waltham, MA USA). Reactions were performed under optimized conditions; denaturation at 95OC for 10 min, cyclic denaturation at 95OC for 10 sec, annealing for 1 minute (Table 2), and extension at 72OC for 1 min. Three reference genes; 18S, ß-actin, and GAPDH were selected for this study, and normalization was done using their mean. Real-time PCR reactions for each gene were performed in triplicate and a mean Cq value was used for fold change calculation. Fold expression was calculated with the 2-ΔΔCq formula.
Results
Immunocytochemistry and immunofluorescence: MCF-7 and T-47D cells showed positive staining for ER and PR and were negative for anti-HER-2 antibodies on both ICC and IF assays (Figures 1 & 2). T-47D had a higher expression of PR than ER compared to MCF-7 which showed an overall similar expression (high expression) of ER and PR. The SK-BR-3 and HCC1954 cells were positive for anti-HER-2 antibody and negative for both ER and PR anti-body (Figures 1 & 2). The expression of HER-2 was higher in SK-BR-3 cells than its expression in HCC1954 cells. The HCC1954 cell also showed a week of staining for ER anti-body on ICC. Additionally, in the culture of HCC1954, a spheroid-like structure was observed throughout the flask (Figure 3). Further, MDA-MB-231 and MDA-MB-468 cells were negative for all three ER, PR, and HER-2 clinical markers. So, based on ER/ PR/HER-2 staining, MCF-7/T-47D belongs to Luminal, SK-BR-3/ HCC1954 belongs to HER-2 positive, and MDA-MB-231/MDAMB-468 belongs to TNBC subtype of breast cancer. However, our study also observed noticeable heterogenous expression of ER, PR, and HER-2 markers in MCF-7, T-47D, SK-BR-3, and HCC1954 cells across the culture plate.
Real-time PCR: On real-time PCR, MCF-7 and T-74D cells had higher expression of the HER-2 gene (MCF-7 log fold 12475; SD±0.25 and T-74D log fold 96433.20; SD±0.006) compared to ESR1 (i.e., ER) (MCF-7 log fold 232.3; SD±0.03 and T-47D log fold 943.69; SD±0.028) and PGR (i.e., PR) genes (MCF-7 log fold 207; SD±0.09 and log fold 3658.96; SD±0.05) relative to in MCF-10A cells (Figure 4). Interestingly, mRNA expression of the HER-2 gene was discordant with ICC and IF results, where both these cell types had negative staining for the HER-2 maker. This might be due to enzymatic degradation of HER-2 mRNA which could not be available for translation into protein. Out of ESR1 and ESR2 both MCF-7 and T-47D cells had higher expression of the ESR1 gene compared to ESR2 (Figure 4). ESR1 and ESR2 encode for nuclear receptors ERα and ERß respectively. ESRα36 and ESRα66 are splice variants of ESR1 those not considered for breast cancer molecular subtyping, though they are tumor promoters. Both these variants are also expressed in breast carcinoma. On evaluation of ESRα36 and ESRα66, both MCF-7 and T47D cells showed higher expression of ESRα66 [(MCF-7, ESRα36 log fold15.08; SD±0.034 vs ESRα66 log fold 3054.26; SD±0.053) and (T-47D, ESRα36 log fold 1098.48 SD ± 0.06 vs ESRα66 log fold 5372.51 SD±0.12)] (Figure 4). Expression of AR was higher in T-47D (log fold 1360.23; SD±0.04) compared to MCF-7 (log fold 954.53; SD±0.15).
In contrast to ICC and IF results, on real-time assay SK-BR-3 had a much higher relative expression of HER-2 gene i.e., log fold 2362800.33; SD±0.04, than in HCC1954 cells i.e., log fold 14161.55; SD±0.09 (Figure 4). However, expression of ERS1 and ERS2 was minimal or basal in both SK-BR-3 and HCC1954 cells which were in concordance with ICC and IF observation. The PGR gene was downregulated in SK-BR-3 and upregulated in HCC1954 cells (log fold 73.54 SD ± 0.03). Whereas SK-BR-3 showed higher expression of ERS1 splice variants ERα36 and ERα66 i.e., log fold 714.17 SD ± 0.16 and log fold 74698.07 SD ± 0.03 respectively (Figure 4). These findings showed that SK-BR-3 and HCC1954 cell lines had similar expression of ER, PR, and HER-2 markers at both mRNA as well as protein levels. Further, the AR gene had less expression in HCC1954 (log fold 141.88; SD±0.33) and SK-BR-3 cells (log fold 440.49 SD ± 0.05) compared to MCF-7 and T-47D cell lines.
Moreover, the expression of clinical markers ER, PR, and HER-2 was very low in MDA-MB-231 and MDA-MB-468 cell lines (Figure 4) which matched with their expression at protein levels determined by ICC and IF (Figures 1 & 2). PGR was observed as downregulated in both MDA-MB-231 and MDA-MB-468 (Figure 4). In addition, ERS1, ESR2, and splice variants of ERS1 (ERα36, ERα66) also showed lower expression in both these cell lines. Therefore, MDA-MB-231 and MDA-MB-468 cell lines represented the phenotype overlapping of breast cancer clinical subtype TNBC. Our study observed less expression of AR in MDAMB-231 and MDA-MB-468 cells compared to its expression in other used cell lines (Figure 4). Clinical TNBC breast cancer samples with positive AR expression called apocrine (a histological subtype). We observed high AR expression in MDA-MB-231, not in MDA-MB-468, therefore MDA-MB-231 might have apocrine histology but require determination of AR at the protein level.
We were also interested in observing the amplification ERBB2 gene, which is routinely done on clinical samples as recommended by the American Society of Clinical Oncology (ASCO)/College of American Pathologists for HER-2 diagnosis. In this study, we have determined EBBR2 gene amplification using real-time PCR. Our study observed ERBB2 gene amplification only in HCC1954 cells (log fold 80.86 SD ± 0.15) compared to the rest of the cell lines tested relative to MCF-10A (Figure 5). Therefore, it shows the overlapping of the HCC1954 cell line with clinical HER-2-positive breast cancer samples.
Discussion
Immortalized cell lines represent a widely used model system in biomedical research due to their ease of handling, authentication, and availability. They offer numerous advantages over primary cells, including cost-effectiveness and circumvention of ethical concerns. Typically, cell lines provide a homogeneous population of cells, ensuring reproducibility in experiments. This model system has significantly advanced biological research and finds applications in antibody/vaccine production, therapeutic protein synthesis, drug testing, and the study of gene expression and function [12,13]. A multitude of diseasespecific cell lines, including those for cancer, are accessible from repositories such as the American Type Culture Collection (ATCC), offering researchers a diverse selection for experimentation. However, despite their utility, careful consideration is warranted when using cell lines as substitutes for primary cells. Genetic modifications and phenotypic changes during continuous passaging can lead to discrepancies between cell lines and primary cells, potentially resulting in heterogeneous cell populations over time. In the context of breast cancer research, cell lines specific to histological and molecular subtypes are available and are utilized to recapitulate in vivo conditions. Nonetheless, the continuous division of cell lines may alter their primary features, necessitating characterization before experimentation. In this study, we characterized a panel of breast cancer cell lines-MCF-7, T-47D, SK-BR-3, HCC1954, MDA-MB-231, MDA-MB-468, and MCF-10A-using clinical markers ER, PR, and HER-2. Analysis of protein and mRNA expression levels revealed heterogeneous marker expression across cell lines, with discrepancies observed between mRNA and protein levels. Immunohistochemistry remains the gold standard for evaluating these markers in clinical breast cancer samples. While our study found consistent protein expression of ER, PR, and HER-2 in cell lines mirroring ATCC records, real-time PCR data revealed variations at the mRNA level relative to protein levels. Discrepancies between mRNA and protein levels could arise from various factors such as transport mechanisms, translational regulation, or differences in transcriptional rates among cells [14,15]. Additionally, we evaluated the expression of ERα36 and HER-2 gene amplification, observing variations among cell lines. Furthermore, we investigated the expression of Androgen Receptor (AR) and its potential impact on breast cancer histology and phenotype and characterized differences in expression levels among cell lines [12-14].
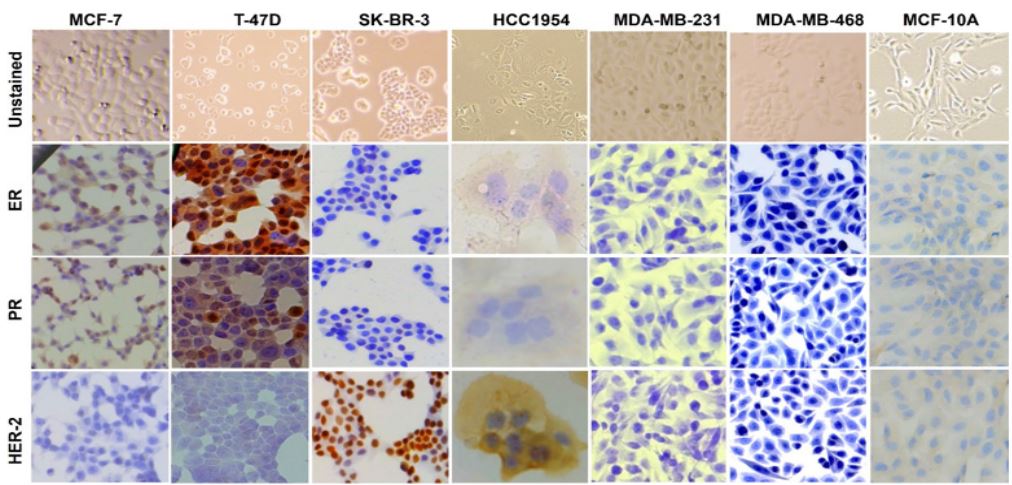
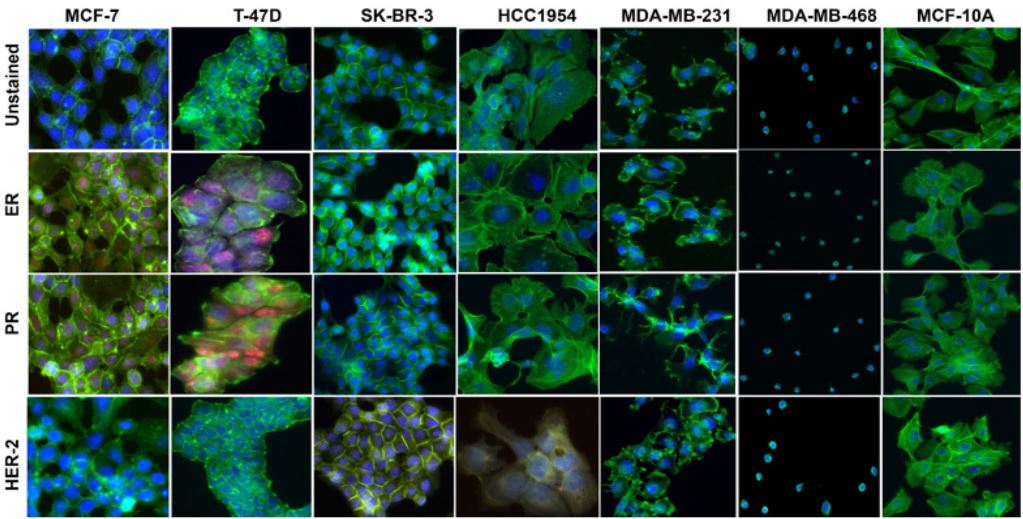

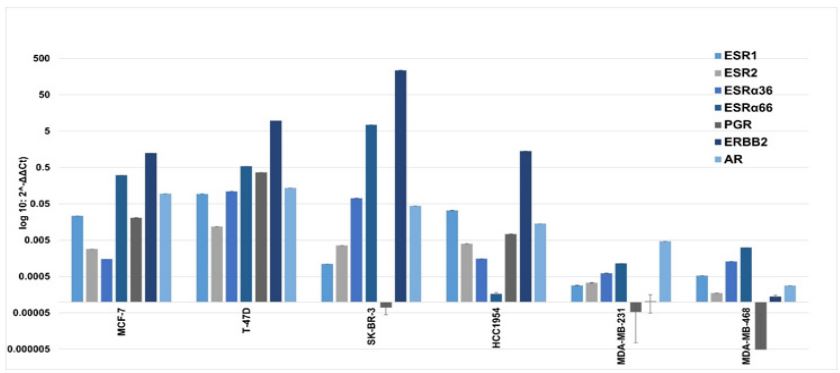
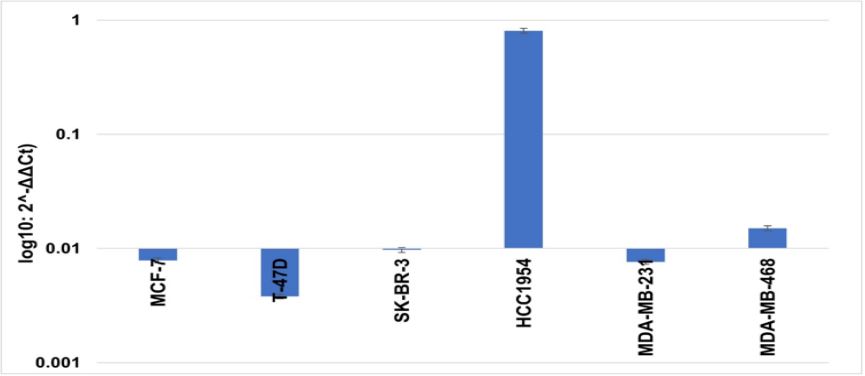
Table 1: Antibodies detail.
| Anti-body | Clonality | Concentration | Dilution |
|---|---|---|---|
| Anti-Estrogen Receptor alpha (SP1) |
Rabbit Monoclonal | 0.014 mg/mL | 1:100 |
| Anti-Progesterone Receptor (SP42) |
Rabbit monoclonal | 0.242 mg/mL | 1:100 |
| Anti-ErbB2/HER-2 (EPR19547-12) |
Rabbit monoclonal | 0.651 mg/mL | 1:100 |
| Secondary, anti-Rabbit HRP |
Goat/IgG polyclonal | 0.8 mg/mL | 1:500 |
| Alexa Flout 555 goat anti-rabbit IgG |
Goat/IgG polyclonal | 2 mg/mL | 1.2:600 |
| Fluorescent phalloidin iFluor 488 |
- | 2 mg/mL | 1:1000 |
Table 2: Primer sequence.
| Gene | Forward (5’-3’) | Reverse (5’-3’) | Annealing |
|---|---|---|---|
| ESR1 | GCTTACTGAC- CAACCTGGCAGA |
GGATCTCTAGCCAGGCA- CATTC |
60OC |
| ESR2 | ATGGAGTCTGGTC- GTGTGAAGG |
TAACACTTCCGAAGTCG- GCAGG |
60OC |
| ESRα36 | CCAAGAATGTTCAAC- CACAACCT |
GCACGGTTCATTAA- CATCTTTCTG |
60OC |
| ESRα66 | AAGAAAGAACAA- CATCAGCAGTAA |
GGGCTATGGCTTGGTTA- AACAT |
60OC |
| PGR | GTCGCCT- TAGAAAGTGCTGT- CAG |
GCTTGGCTTTCATTTG- GAACGCC |
60OC |
| ERB2 | GGAAGTACACGAT- GCGGAGACT |
ACCTTCCTCAGCTCC- GTCTCTT |
60OC |
| ß-ACTIN | CACCATTGGCAAT- GAGCGGTTC |
AGGTCTTTGCGGATGTC- CACGT |
60OC |
| GAPDH | GTCTCCTCT- GACTTCAACAGCG |
ACCACCCTGTTGCTGTAGC- CAA |
60OC |
| 18S | ACCCGTT- GAACCCCATTCGTGA |
GCCTCACTAAACCATC- CAATCGG |
60OC |
| ERBB2_ GN* |
GCACAGGGT- GGGCCTAGTC |
CTTGCACACTGCAGGTT- TAAC |
60OC |
| RPP30 | CAGATGTTGGGTAC- TAATGAC |
CCAGGTATCTTCAGGTA- AAGTG |
60OC |
| *For ERBB2 gene amplification | |||
Table 3: Comparing the expression status of ER, PR, and HER-2 markers at protein and mRNA levels.
| Cell line | MCF-7 | T-47D | SK-BR-3 | HCC1954 | MDA-MB-231 | MDA-MB-468 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein | mRNA | Protein | mRNA | Protein | mRNA | Protein | mRNA | Protein | mRNA | Protein | mRNA | |
| ER | + | + | + | + | - | + | - | + | - | + # | - | + # |
| PR | + | + | + | + | - | + | - | + | - | - | - | - |
| HER-2 ↑ | - | + ↑ | - | + ↑ | + | + | + | + ↑ | - | + # | - | + # |
Note: = Higher expression; # = Low expression
Table 4: Summarizing the resemblance of each cell line with intrinsic breast cancer subtypes and proposing their best suitability.
| Cell line | Intrinsic breast cancer subtype | Confirmed by | Suitability |
|---|---|---|---|
| MCF-7 | Luminal subtype | At protein as well as mRNA level | Best suitable |
| SK-BR-3 | Her-2 positive/enriched | At protein as well as mRNA level | Best suitable |
| MDA-MB-231 | TNBC | At protein as well as mRNA level | Best suitable |
| MDA-MB-468 | TNBC | At protein as well as mRNA level | Best suitable |
| T-47D | Luminal subtype | At the protein level only, not at the mRNA level | Not well suitable |
| HCC1954 | Her-2 positive/enriched | At the protein level only, not at the mRNA level | Not well suitable |
The HCC1954 cell line exhibited the formation of spheroidlike structures when cultured in two-Dimensional (2D) conditions utilizing basic growth media devoid of specialized supplements. This observation is noteworthy given that previous literature has documented the propensity of HCC1954 cells to adopt spheroid configurations in both 2D and three-Dimensional (3D) cultures, albeit typically contingent upon the addition of specific supplements [15-17]. The emergence of spheroid-like structures within cell cultures holds considerable importance for multiple reasons, including the facilitation of a more accurate mimicry of tissue architecture and physiology. This attribute renders such cultures particularly advantageous for applications in drug testing and development, stem cell research, and the modeling of diseases. Consequently, the inherent capacity of HCC1954 to form spheroids under minimalistic culture conditions without the necessity for specialized supplements may position it as a preferred cell line for experiments aimed at exploring these areas.
This study delineated a precise correlation between the MCF-7, SK-BR-3, and MDA-MB-231/MDA-MB-468 cell lines and the Luminal, HER-2 positive/enriched, and Triple-Negative Breast Cancer (TNBC) subtypes, respectively. These cell lines exhibited specific expression patterns of Estrogen Receptor (ER), Progesterone Receptor (PR), and Human Epidermal Growth Factor Receptor 2 (HER-2) markers, aligning with the molecular subtypes of breast cancer at both protein and mRNA levels, as presented in Table 4. Conversely, the T-47D and HCC1954 cell lines did not exhibit a direct correspondence with the clinical molecular subtypes of breast cancer, attributable to the discordant expression of ER, PR, and HER-2 markers at the protein and mRNA levels (Table 4). This observation contrasts with prior studies, which reported a concordance rate exceeding 90% for ER, PR, and HER-2 markers between immunohistochemistry (IHC) and real-time PCR analyses in clinical samples, particularly regarding the HER-2 gene [18]. This high degree of concordance has facilitated the development of several real-time PCR-based commercial kits, including PAM50, MammaPrint, TargetPrint, and BluePrint, designed for the molecular subtyping of breast cancer tissue samples [19,20].
Thus, our study underscores the importance of evaluating protein expression of clinical markers for breast cancer subtyping in cell lines, as mRNA levels reflect diverse phenotypes.
Conclusion
In conclusion, our study utilized ICC, IF, and real-time assays to characterize the expression profiles of clinical markers ER, PR, and HER-2 in a panel of breast cancer cell lines. Our findings indicate that MCF-7 and T-47D cell lines exhibit characteristics consistent with the Luminal subtype, while SK-BR-3 and HCC1954 cell lines are indicative of the HER-2 enriched subtype. Conversely, MDA-MB-231 and MDA-MB-468 cell lines resemble the TNBC subtype. Notably, significant discrepancies were observed between protein and mRNA levels of these markers, highlighting the need for comprehensive characterization methods. Based on our analysis, MCF-7, SK-BR-3, MDA-MB-231, and MDA-MB-468 cell lines are identified as ideal models for studying breast cancer molecular subtypes.
Declarations
Funding: Brown Center for Immunotherapy/ Indiana University School of Medicine
Conflict of interest: Authors declared no conflict of interest.
Acknowledgment: The authors would like to acknowledge the Brown Center for Immunotherapy/ Indiana University School of Medicine, Indiana University, Indiana, USA-46202 for supporting the study.
References
- Wilkinson L, Gathani T. Understanding breast cancer as a global health concern. Br J Radiol. 2022; 95(1130): 20211033. doi: 10.1259/bjr.20211033.
- Reis-Filho JS, Pusztai L. Gene expression profiling in breast cancer: classification, prognostication, and prediction. Lancet. 2011 Nov 19; 378(9805): 1812-23. doi: 10.1016/S0140-6736(11)61539-0.
- Blows FM, Driver KE, Schmidt MK, Broeks A, van Leeuwen FE, et al. Subtyping of breast cancer by immunohistochemistry to investigate a relationship between subtype and short and long term survival: A collaborative analysis of data for 10,159 cases from 12 studies. PLoS Med. 2010; 7(5): e1000279. doi: 10.1371/journal.pmed.1000279.
- Lüönd F, Tiede S, Christofori G. Breast cancer as an example of tumour heterogeneity and tumour cell plasticity during malignant progression. Br J Cancer. 2021; 125(2): 164-175. doi: 10.1038/s41416-021-01328-7.
- Riaz M, van Jaarsveld MT, Hollestelle A, Prager-van der Smissen WJ, Heine AA, et al. miRNA expression profiling of 51 human breast cancer cell lines reveals subtype and driver mutation-specific miRNAs. Breast Cancer Res. 2013; 15(2): R33. doi: 10.1186/bcr3415.
- Cope LM, Fackler MJ, Lopez-Bujanda Z, Wolff AC, Visvanathan K, et al. Do breast cancer cell lines provide a relevant model of the patient tumor methylome? PLoS One. 2014; 9(8): e105545. doi: 10.1371/journal.pone.0105545.
- Elstrodt F, Hollestelle A, Nagel JH, Gorin M, Wasielewski M, et al. BRCA1 mutation analysis of 41 human breast cancer cell lines reveals three new deleterious mutants. Cancer Res. 2006; 66(1):41-5. doi: 10.1158/0008-5472.CAN-05-2853.
- Kao J, Salari K, Bocanegra M, Choi YL, Girard L, et al. Molecular profiling of breast cancer cell lines defines relevant tumor models and provides a resource for cancer gene discovery. PLoS One. 2009; 4(7): e6146. doi: 10.1371/journal.pone.0006146.
- Neve RM, Chin K, Fridlyand J, Yeh J, Baehner FL, et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell. 2006; 10(6): 515-27. doi: 10.1016/j.ccr.2006.10.008.
- Charafe-Jauffret E, Ginestier C, Monville F, Finetti P, Adélaïde J, et al. Gene expression profiling of breast cell lines identifies potential new basal markers. Oncogene. 2006; 25(15): 2273-84. doi: 10.1038/sj.onc.1209254.
- Lehmann BD, Bauer JA, Chen X, Sanders ME, Chakravarthy AB, et al. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J Clin Invest. 2011; 121(7): 2750-67. doi: 10.1172/JCI45014.
- Gómez-Lechón MJ, Donato MT, Castell JV, Jover R. Human hepatocytes as a tool for studying toxicity and drug metabolism. Curr Drug Metab. 2003; 4(4): 292-312. doi: 10.2174/1389200033489424.
- MacDonald C. Development of new cell lines for animal cell biotechnology. Crit Rev Biotechnol. 1990; 10(2): 155-78. doi: 10.3109/07388559009068265.
- Titlow JS, Kiourlappou M, Palanca A, Lee JY, Gala DS, et al. Systematic analysis of YFP traps reveals common mRNA/protein discordance in neural tissues. J Cell Biol. 2023; 222(6): e202205129. doi: 10.1083/jcb.202205129.
- Jahin I, Phillips T, Marcotti S, Gorey MA, Cox S, et al. Extracellular matrix stiffness activates mechanosensitive signals but limits breast cancer cell spheroid proliferation and invasion. Front Cell Dev Biol. 2023; 11: 1292775. doi: 10.3389/fcell.2023.1292775.
- Breslin S, O’Driscoll L. The relevance of using 3D cell cultures, in addition to 2D monolayer cultures, when evaluating breast cancer drug sensitivity and resistance. Oncotarget. 2016; 7(29): 45745-45756. doi: 10.18632/oncotarget.9935.
- Franchi-Mendes T, Lopes N, Brito C. Heterotypic Tumor Spheroids in Agitation-Based Cultures: A Scaffold-Free Cell Model That Sustains Long-Term Survival of Endothelial Cells. Front Bioeng Biotechnol. 2021; 9: 649949. doi: 10.3389/fbioe.2021.649949.
- Badve SS, Baehner FL, Gray RP, Childs BH, Maddala T, et al. Estrogen- and progesterone-receptor status in ECOG 2197: comparison of immunohistochemistry by local and central laboratories and quantitative reverse transcription polymerase chain reaction by central laboratory. J Clin Oncol. 2008; 26(15): 2473-81. doi: 10.1200/JCO.2007.13.6424. Erratum in: J Clin Oncol. 2008; 26(20): 3472.
- Veerla S, Hohmann L, Nacer DF, Vallon-Christersson J, Staaf J. Perturbation and stability of PAM50 subtyping in populationbased primary invasive breast cancer. NPJ Breast Cancer. 2023; 9(1): 83. doi: 10.1038/s41523-023-00589-0.
- Nguyen B, Cusumano PG, Deck K, Kerlin D, Garcia AA, et al. Comparison of molecular subtyping with Blue Print, Mamma Print, and Target Print to local clinical subtyping in breast cancer patients. Ann Surg Oncol. 2012; 19(10): 3257-63. doi: 10.1245/s10434-012-2561-6.